

Published on May 29, 2026
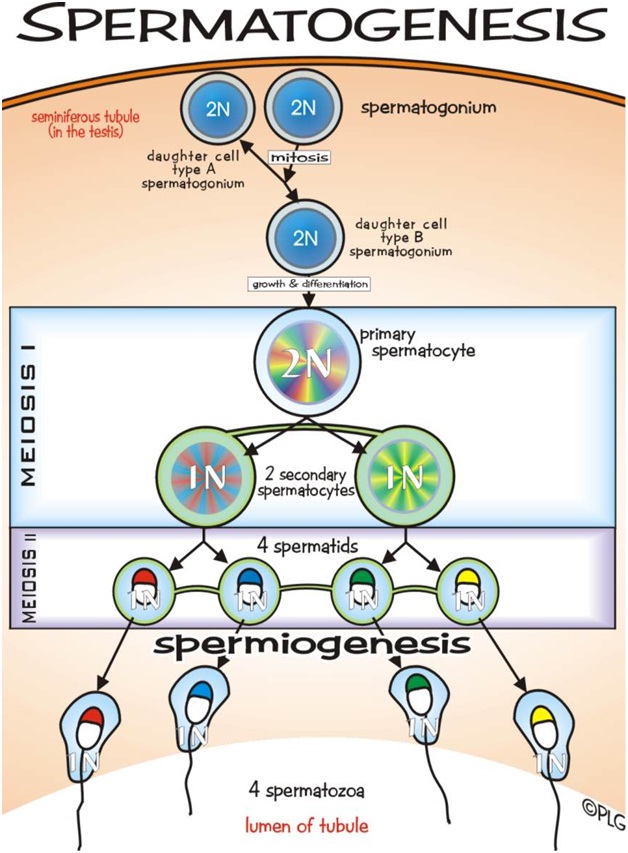
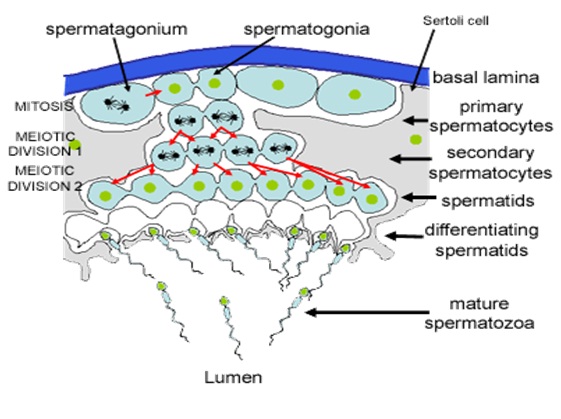
Spermatogenesis is the process in which spematozoa are produced from male primordial germ cells by way of mitosis and meiosis. The initial cells in this pathway are called spematogonia, which yield primary spermatocytes by mitosis. The primary spermatocyte divides meiotically into two secondary spermatocytes; each secondary spermatocyte then completes meiosis as it divides into two spermatids. These develop into mature spermatozoa, also known as sperm cells. Thus, the primary spermatocyte gives rise to two cells, the secondary spermatocytes, and the two secondary spermatocytes by their subdivision produce four spermatozoa.
Spermatozoa are the mature male gametes in many sexually reproducing organisms. Thus, spermatogenesis is the male version of gametogenesis. In mammals it occurs in the male testes and epididymis in a stepwise fashion. Spermatogenesis is highly dependent upon optimal conditions for the process to occur correctly, and is essential for sexual reproduction. DNA methylation and histone modification have been implicated in the regulation of this process. It starts at puberty and usually continues uninterrupted until death, although a slight decrease can be discerned in the quantity of produced sperm with increase in age.
Spermatogenesis produces mature male gametes, commonly called sperm but specifically known as spermatozoa, which are able to fertilize the counterpart female gamete, the oocyte, during conception to produce a single-celled individual known as a zygote. This is the cornerstone of sexual reproduction and involves the two gametes both contributing half the normal set of haploid chromosomes to result in a chromosomally normal diploid zygote. To preserve the number of chromosomes in the offspring – which differs between species – each gamete must have half the usual number of chromosomes present in other body cells. Otherwise, the offspring will have twice the normal number of chromosomes, and serious abnormalities may result. In humans, chromosomal abnormalities arising from incorrect spermatogenesis can result in Down syndrome, Klinefelter’s syndrome, and spontaneous abortion
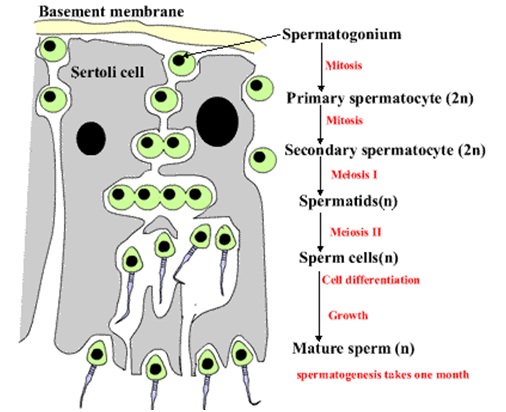
Spermatogenesis takes place within several structures of the male reproductive system. The initial stages occur within the testes and progress to the epididymis where the developing gametes mature and are stored until ejaculation. The seminiferous tubules of the testes are the starting point for the process, where stem cells adjacent to the inner tubule wall divide in a centripetal direction—beginning at the walls and proceeding into the innermost part, or lumen—to produce immature sperm. Maturation occurs in the epididymis.
For humans, entire process of spermatogenesis takes 74 days. Including the transport on ductal system, it takes 3 months. Testes produce 200 to 300 million spermatozoa daily.
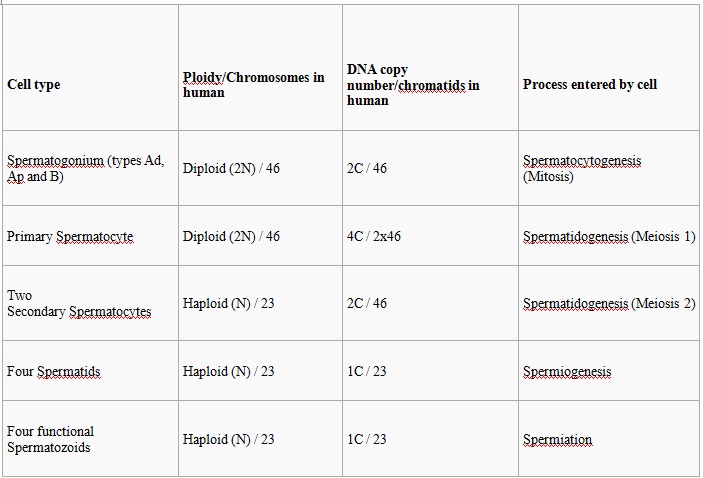
The entire process of spermatogenesis can be broken up into several distinct stages, each corresponding to a particular type of cell in human. In the following table, ploidy, copy number and chromosome/chromatid counts are for one cell, generally prior to DNA synthesis and division (in G1 if applicable). The primary spermatocyte is arrested after DNA synthesis and prior to division.
The process of spermatogenesis as the cells progress from primary spermatocytes, to secondary spermatocytes, to spermatids, to sperm.
Spermatocytogenesis is the male form of gametocytogenesis and results in the formation of spermatocytes possessing half the normal complement of genetic material. In spermatocytogenesis, a diploid spermatogonium which resides in the basal compartment of seminiferous tubules, divides mitotically to produce two diploid intermediate cells called primary spermatocytes. Each primary spermatocyte then moves into the adluminal compartment of the seminiferous tubules and duplicates its DNA and subsequently undergoes meiosis I to produce two haploid seconadry spermatocytes , which will later divide once more into haploid spermatids. This division implicates sources of genetic variation, such as random inclusion of either parental chromosomes, and chromosomal crossover, to increase the genetic variability of the gamete.
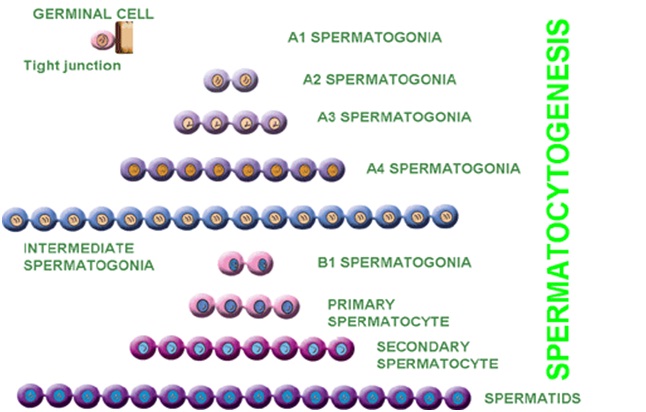
Each cell division from a spermatogonium to a spermatid is incomplete; the cells remain connected to one another by bridges of cytoplasm to allow synchronous development. It should also be noted that not all spermatogonia divide to produce spermatocytes, otherwise the supply would run out. Instead, certain types of spermatogonia divide to produce copies of themselves, thereby ensuring a constant supply of gametogonia to fuel spermatogenesis.
During spermiogenesis, the spermatids begin to grow a tail, and develop a thickened mid-piece, where the microtubules gather and form an axoneme . Spermatid DNA also undergoes packaging, becoming highly condensed. The DNA is packaged firstly with specific nuclear basic proteins, which are subsequently replaced with protamines during spermatid elongation. The resultant tightly packed chromatin is transcriptionally inactive. The golgi apparatus surrounds the now condensed nucleus, becoming the acrosome. One of the centrioles of the cell elongates to become the tail of the sperm.
Maturation then takes place under the influence of testosterone, which removes the remaining unnecessary cytosome and organelles. The excess cytoplasm, known as residual bodies, is phagocytosed by surrounding Sertoli cells in the testes. The resulting spermatozoa are now mature but lack motility, rendering them sterile. The mature spermatozoa are released from the protective sertoli cells into the lumen of the seminiferous tubules in a process called spermiation.
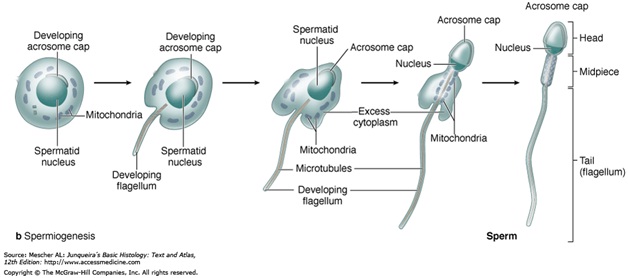
The non-motile spermatozoa are transported to the epididymis in testicular fluid secreted by the Sertoli cells with the aid of peristaltic contraction. While in the epididymis the spermatozoa gain motility and become capable of fertilization. However, transport of the mature spermatozoa through the remainder of the male reproductive system is achieved via muscle contraction rather than the spermatozoon's recently acquired motility.
At all stages of differentiation, the spermatogenic cells are in close contact with Sertoli cells which are thought to provide structural and metabolic support to the developing sperm cells. A single Sertoli cell extends from the basement membrane to the lumen of the seminiferous tubule, although the cytoplasmic processes are difficult to distinguish at the light microscopic level.
Sertoli cells serve a number of functions during spermatogenesis, they support the developing gametes in the following ways:
• Maintain the environment necessary for development and maturation, via the blood testes barrier
• Secrete substances initiating meiosis
• Secrete supporting testicular fluid
• Secrete androgen binding protien (ABP), which concentrates testosterone in close proximity to the developing gametes
• Testosterone is needed in very high quantities for maintenance of the reproductive tract, and ABP allows a much higher level of fertility
• Secrete hormones affecting pituitary gland control of spermatogenesis, particularly the polypeptide hormone, inhibin
• Phagocytose residual cytoplasm left over from spermiogenesis
• They release antimullerian harmone which prevents formation of the Müllerian Duct / Oviduct.
• Protect spermatids from the immune system of the male, via the blood testes barrier
The intercellular adhesion molecules ICAM -1 and soluble ICAM-1 have antagonistic effects on the tight junctions forming the blood-testis barrier. ICAM -2 molecules regulate spermatid adhesion on the apical side of the barrier (towards the LUMEN) during spermatogenesis, the cells are closely associated with sertoli cells which lies at regular interval alone the seminiferous tubules. sertoli cells perform the following tasks:
• are target cells for follicular stimulating hormones (FSH)
• synthesize an androgen binding protein that maintain a high levels of testosterone inside the seminiferous tubules
• maintain the blood-testes barrier which protect the body's immune system from destroying the developing sperm cells
• create an environment that is necessary in the differentiation of sperm cell
• degrade the residual cytoplasm that is shed during spermatogenesis.
The process of spermatogenesis is highly sensitive to fluctuations in the environment, particularly harmones and temperature. Testosterone is required in large local concentrations to maintain the process, which is achieved via the binding of testosterone by androgen binding protein present in the seminiferous tubules. Testosterone is produced by interstitial cells, also known as leydig cells, which reside adjacent to the seminiferous tubules.
Seminiferous epithelium is sensitive to elevated temperature in humans and some other species, and will be adversely affected by temperatures as high as normal body temperature. Consequently, the testes are located outside the body in a sack of skin called the scrotum. The optimal temperature is maintained at 2 Degree Celcius–8 °C (mouse) below body temperature. This is achieved by regulation of blood flow and positioning towards and away from the heat of the body by the cremastaric muscle and the dartos smooth muscle in the scrotum.
Dietary deficiencies (such as vitamins B, E and A), anabolic steroids, metals (cadmium and lead), x-ray exposure, dioxin, alcohol, and infectious diseases will also adversely affect the rate of spermatogenesis.In addition, the male germ line is susceptible to DNA damage caused by oxidative stress, and this damage likely has a significant impact on fertilization and pregnancy. Exposure to pesticides also affects spermatogenesis.
Hormonal control of spermatogenesis varies among species. In humans the mechanism is not completely understood, however it is known that initiation of spermatogenesis occurs at puberty due to the interaction of the hypothalamus, pituitary gland and Leydig cells. If the pituitary gland is removed, spermatogenesis can still be initiated by follicle stimulating harmone and testesterone.
Follicle stimulating hormone stimulates both the production of androgen binding protein by Sertoli cells, and the formation of the blood testis barrier. Androgen binding protein is essential to concentrating testosterone in levels high enough to initiate and maintain spermatogenesis, which can be 20–50 times higher than the concentration found in blood. Follicle stimulating hormone may initiate the sequestering of testosterone in the testes, but once developed only testosterone is required to maintain spermatogenesis. However, increasing the levels of follicle stimulating hormone will increase the production of spermatozoa by preventing the apoptysis of type A spermatogonia. The hormone inhibin acts to decrease the levels of follicle stimulating hormone. Studies from rodent models suggest that gonadotropin hormones (both LH and FSH) support the process of spermatogenesis by suppressing the proapoptotic signals and therefore promote spermatogenic cell survival.
The Sertoli cells themselves mediate parts of spermatogenesis through hormone production. They are capable of producing the hormones estradiol and inhibin. The Leydig cells are also capable of producing estradiol in addition to their main product testosterone.
1. www.merriam-webster.com/dictionary/gametogenesis
2. https://www.landesbioscience.com/journals/spermatogenesis/
3. www.sparknotes.com/testprep/books/sat2/.../chapter7section2.rhtm
4. www.embryology.ch/anglais/cgametogen/oogenese01.html
5. www.ncbi.nlm.nih.gov › NCBI › Literature › Bookshelf